
触覚パターン認知の神経回路演算原理を光遺伝学により解明
‐ラット大脳皮質バレル野における多点入力間相互作用の光遺伝学的研究
東京大学物性研究所 八尾 寛
Liu Y, Ohshiro T, Sakuragi S, Koizumi K, Mushiake H, Ishizuka T, Yawo H.
Optogenetic study of the response interaction among multi-afferent inputs in the barrel cortex of rats.
Sci Rep., 9(1): 3917, 2019年. DOI: 10.1038/s41598-019-40688-2.
https://www.nature.com/articles/s41598-019-40688-2
ヒトは触覚などの体性感覚を介して、外界のさまざまな情報をえています。たとえば、皮膚の各部位に与えられた接触、圧力、振動などの情報は、それぞれの感覚受容器に神経を送っているメカノ受容ニューロンを経て、大脳皮質体性感覚野へ送られます。感覚受容器の部位と大脳皮質部位が対応するように情報が送られるので、体の部位と対応した知覚が生じます。このようなトポグラフィカル知覚に加え、私たちは触覚を介して、ものの形、大きさ、運動、手触りなどの複合的な知覚を得ています。しかし、これら複合的な知覚が、脳の中でどのように作り出されているかに関する神経生理学的研究は、ほとんどなされていません。これは、動物実験において、皮膚をパターン刺激することが難しいという技術的なハードルが存在するためです。
マウスやラットなどの夜行性哺乳類には,顔面両側に発達した頬ひげ(ウィスカ)があり,空間認知に関わる重要な感覚器になっています(図1A).約30本のウィスカは,口先に規則正しく配置され,A1, A2, …, B1, B2, …などの番地が振り当てられています(図1B)。各々のウィスカの接触は,毛根において感知され,三叉神経→脳幹→視床を経て,反対側の大脳皮質一次感覚野へトポグラフィカルに情報が受け渡されます.一次感覚野においては,ウィスカの2次元的な配列に対応した2次元的な樽表面構造(バレル)が形成されているので、この脳領域はバレル野とよばれています.すなわち,ウィスカに接触した物体の形や運動などの時空間パターン情報は,バレル野の時空間的活動パターンに反映されます.ウィスカ―バレル野は,体性感覚におけるパターン認識やその発達・可塑性の優れたモデルになっています.
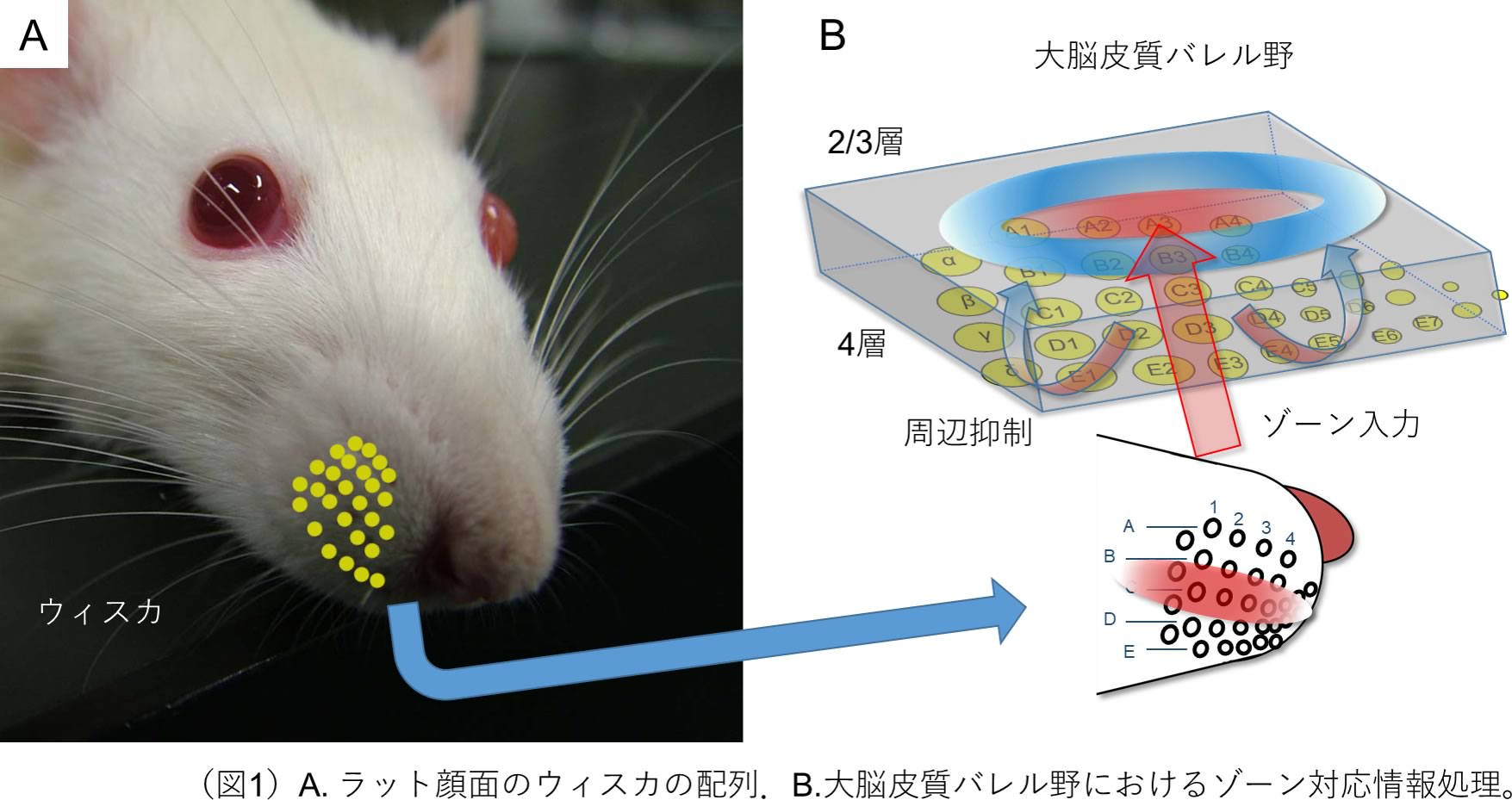
筆者の研究グループおよびカール・ダイセロース研究グループは、それぞれ独立に、単細胞緑藻類クラミドモナスの光受容タンパク質の一つ、チャネルロドプシン2 (ChR2)をニューロンに作らせることにより、ニューロン活動を光操作できることを、世界に先駆けて報告しました(八尾・石塚、特願2005-34529)。この技術は、光遺伝学(オプトジェネティクス)とよばれ、神経生理学研究のブレークスルーになりました。筆者のグループは、さらに、触覚をつかさどるメカノ受容ニューロン特異的にChR2遺伝子を組込んだトランスジェニックラット(W-TChR2V4)を作製しました。W-TChR2V4ラットの皮膚にある感覚受容器に入力している感覚神経末端部にもChR2が発現しているので、このラットは、皮膚に照射した青色光を触覚として知覚します。特にウィスカの毛根周囲に分布するメカノ受容神経終末にChR2が高密度に分布し、反対側の大脳皮質バレル野において、ウィスカ光刺激に対する神経活動応答が引き起こされました。そこで、W-TChR2V4ラットのウィスカ毛根の光刺激により、触覚の時間・空間的なパターンを作り出し、パターン入力が大脳皮質において、どのように情報処理されるかの研究を着想しました。具体的には、光源(青色LED)と4×4に配列している16本のウィスカの毛根をそれぞれ光ファイバーでつなぎ、コンピューターで作成した時系列パターンで、個々の光源を独立にオン・オフすることにより、時空間刺激パターンをさまざまに与えながら、大脳皮質浅層(2/3層)応答を計測し、入出力関係から、その演算原理を解明しました。実験には、本支援(生理機能解析支援/多機能集積化電極等による多次元生理機能解析支援)を受け、ポリ尿素絶縁膜タングステン電極を用いました。その結果、大脳皮質の電気活動を計測するにあたり、良好な信号/ノイズ比が得られ、精度が向上しました。本研究では、ウィスカ光刺激直後、最初に計測される大脳皮質応答にフォーカスしました。
4か所のウィスカの組み合わせは、16C4 (=1820)通りあります。すなわち、一つのウィスカあたり15C3 (=455)通りのパターンがあります。各ウィスカを一つずつ光刺激したときに比べ、組み合わせることにより大脳皮質応答確率が小さくなる傾向があることに気づきました。そこで、大脳皮質応答確率が複数のウィスカ入力の相互作用の関数であると仮定し、入出力関係を多変量解析しました。その結果、複数のウィスカ入力を受け取る大脳皮質ニューロンが多くあり、そのほとんどが水平方向の刺激パターンに鋭敏であることがわかりました。また、その周囲のウィスカ入力が抑制的にはたらいていることも確かめられました(図1B)。すなわち、ある水平ゾーンの入力は、その上下レベルのゾーン入力と対比的に識別されていました。
ウィスカとパレル野の間には、古典的なトポグラフィカルな情報と本研究で見出されたゾーン対応情報が並列的に受け渡しされていると考えられます。ゾーン対応情報は、ウィスカで受け取った情報を、いち早く大脳皮質に届ける機能に最適化しています。たとえば、ウィスカが接触した対象の位置や大きさを大まかにとらえ、行動で素早く対応する機能を担っているのかもしれません。これに対し、トポグラフィカルな情報は、対象表面の凹凸や運動にともなうウィスカの曲げを検知し、その詳細を解析する機能に最適化しているのではないでしょうか。両者を組み合わせることにより、すばやくかつ正確な情報処理が実現されていると考えています。
