
piRNA生合成機構「ピンポン経路」の解析に有用なショウジョウバエ培養細胞株の作出
東京大学大学院理学系研究科生物科学専攻 塩見美喜子
Sumiyoshi, T., Sato, K., Yamamoto, H., Iwasaki, YW., Siomi, H., Siomi, MC.
Loss of l(3)mbt leads to acquisition of the ping-pong cycle in Drosophila ovarian somatic cells.
Genes Dev. 30: 1617-1622 DOI: 10.1101/gad.283929.116. (2016)
http://genesdev.cshlp.org/content/30/14/1617.long
PIWI-interacting RNA (piRNA)は、生殖細胞特異的に産生される24~30塩基長の小分子RNAであり、PIWIタンパク質ファミリーと複合体を形成することで機能する。piRNAの多くはトランスポゾンの相補的な配列を有し、塩基配列特異的にPIWI-piRNA複合体がトランスポゾンRNAに作用することで、それらの発現を抑制し、生殖細胞のゲノムの品質管理を担う。piRNAが産生されないとトランスポゾンによって生殖ゲノムが損傷し、卵や精子の形成不全を引き起こす。
piRNAの産生経路に関しては3つのモデル、プライマリー経路およびピンポン経路、フェイジング経路が提唱されている。ショウジョウバエ卵巣は、主に生殖細胞とそれを取り囲む濾胞細胞から構成され、プライマリー経路とフェイジング経路は両細胞でみられるが、ピンポン経路は生殖細胞でのみ観察される(図1A)。そのため、ピンポン経路によるpiRNA生合成機構を研究するには、卵巣や精巣といった生殖組織を使わざるを得ない。生殖組織を用いて生化学的な解析を行う上で問題となるのは、生殖組織は生殖細胞以外の細胞が混じったヘテロな細胞集団であり、それが解析上のノイズとなり得ること、さらに、個体数の制限や組織解剖の手間といったサンプル調整の困難さ、である。これらの点は、ショウジョウバエに限らず、マウスなどのモデル生物であってもpiRNA研究を推進する上で律速となっていた。そのため、生化学的な解析には組織よりも培養細胞が適しており、均一な細胞集団を用いたシンプルな実験系での解析を可能とする。つまり、ピンポン経路を有する培養細胞の作出が、本題の解消につながると考えられた。
これまでに、ショウジョウバエのがん遺伝子lethal (3) malignant brain tumor [l(3)mbt]を機能欠損させると脳腫瘍が生じ、その腫瘍細胞ではVasaやAub、AGO3といった生殖細胞特異的なpiRNA生合成因子が異所的に発現することが報告されていた。そこで、我々は、l(3)mbt遺伝子に着目し、ショウジョウバエ濾胞細胞由来の培養細胞株OSCにおいて、l(3)mbt遺伝子をノックアウトした培養細胞株「Δmbt-OSC」をCRISPR-Cas9システムを用いて作出した(図1A)。Δmbt-OSCでは、本来OSCでは発現していないVasaやAub、AGO3などの生殖細胞特異的piRNA生合成因子が異所的に発現し(図1B)、さらに、ピンポン経路を介したpiRNAの産生も起こっていたことから(図1C)、ピンポン経路をはじめとした、生殖細胞でのpiRN A生合成経路を生化学的に分子レベルで解明できる優れた研究ツールであると考えられる。Δmbt-OSCの培養細胞としての性状などは他のショウジョウバエ培養細胞株と変わらず、容易に継代・維持が可能である。現在、Δmbt-OSCは、Drosophila Genomics Resource Center (DGRC) でも維持されており、そちらからも入手可能である(https://dgrc.bio.indiana.edu/Home)。
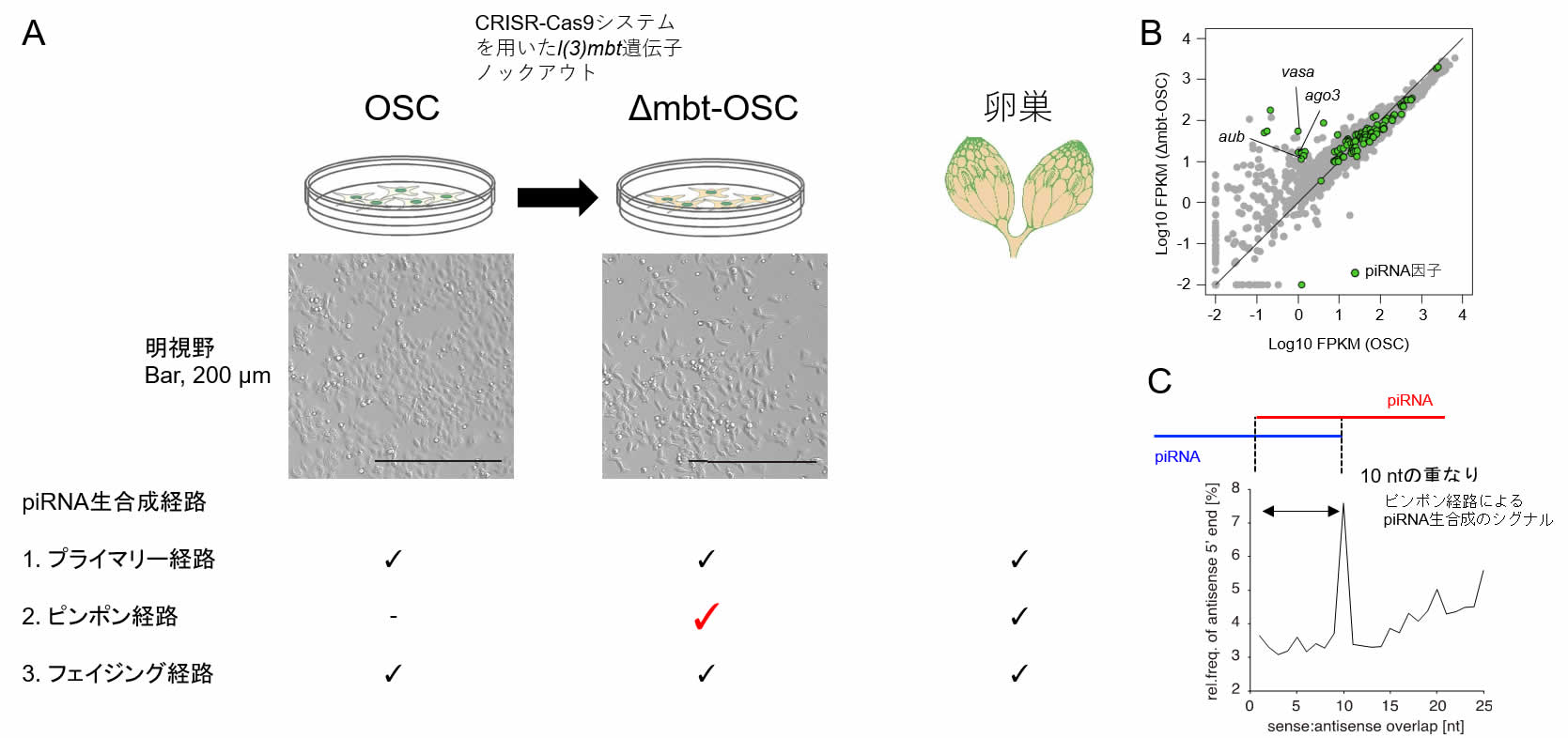
図1. CRISPR-Cas9システムを用いたショウジョウバエ培養細胞株「Δmbt-OSC」の作出.
A. Δmbt-OSCではピンポンpiRNA生合成経路が活性化する.
B. RNA-seqによるΔmbt-OSCにおける生殖細胞特異的piRNA因子(Aub, AGO3, Vasaなど)の異所的な発現.
C. Δmbt-OSCにおけるピンポン経路を示唆するシグナル(ピンポンシグナル、piRNA間の10塩基の重なり)の検出.
![]()
