線虫の体はどのようにできるの?(発生)
線虫の体はどのようにできるの?(発生)
目次
Illustrated by Hiroko Uchida
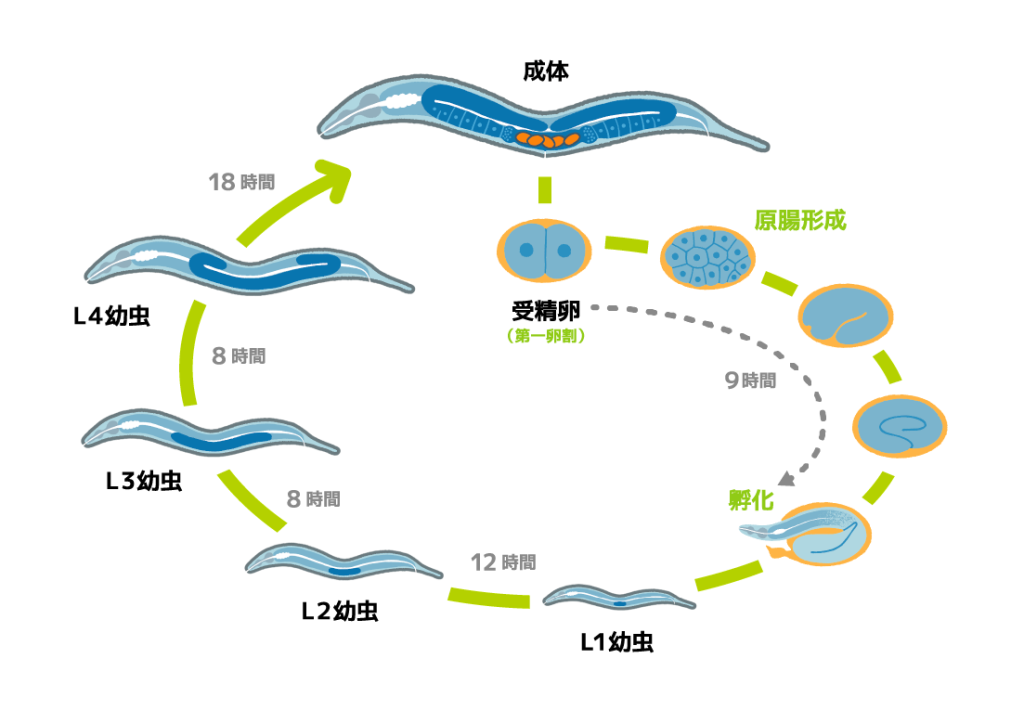
私たちヒトと同じように、線虫もたった一つの細胞である受精卵が分裂を繰り返し、多数の細胞からなる体ができあがります。線虫は体が透明なため、体ができあがる過程を顕微鏡で詳細に観察できます。受精後たった9時間で孵化し、その後、わずか3日間で成虫にまで成長します。さらには、細胞系譜(細胞分裂の順番や分裂した細胞がどの器官を形成するかという細胞運命)があらかじめ決まっているという特徴もあり、体ができあがる過程を研究するのにうってつけの生き物です。実際に、体ができあがるさまざまな仕組みが線虫を使って明らかにされてきて、その多くはヒトにも共通すると考えられています。
監修者
木村 暁
東京大学大学院博士課程修了。現在、国立遺伝学研究所教授として「細胞が建築される仕組み」の研究を担当している。
研究室HP
https://www.nig.ac.jp/nig/ja/research/organization-top/laboratories/kimura
©2026 虫の集い(線虫研究者コミュニティ)「序論」CC BY
1. 減数分裂(生殖腺で精子と卵子ができるまで)
精子と卵子を作り出す特別な細胞分裂:減数分裂
減数分裂は、精子、卵子などの生殖細胞を生み出す特別な細胞分裂です。細胞が正常に働くためには、適切な数の染色体を持っていることが重要であり、例えばヒトの場合は、母方から23本、父方から23本の染色体を受け継ぐことで、46本の染色体を持っています。染色体の数が多すぎても少なすぎても細胞の働きに異常をきたすため、正しい数と組み合わせ(1番染色体から22番染色体からなる常染色体と、性染色体)の染色体を親から受け継ぐことが生物にとって重要です。
減数分裂では、元となる細胞(ヒトの場合は46本の染色体を持つ細胞)から、染色体を正確に半数化した生殖細胞(ヒトの場合は23本の染色体を持つ精子・卵子)を生み出します。この精子と卵子が受精により合体することで、ヒトの場合は再び46本の染色体を持つ1細胞が誕生し、そこから子孫となる個体が誕生します。減数分裂におけるエラーは、人間の場合、異常な精子や卵子の産出、そして不妊、流産や染色体異常症などにつながります。
減数分裂のしくみ
では減数分裂において、染色体はどのように半数化されているのでしょうか?
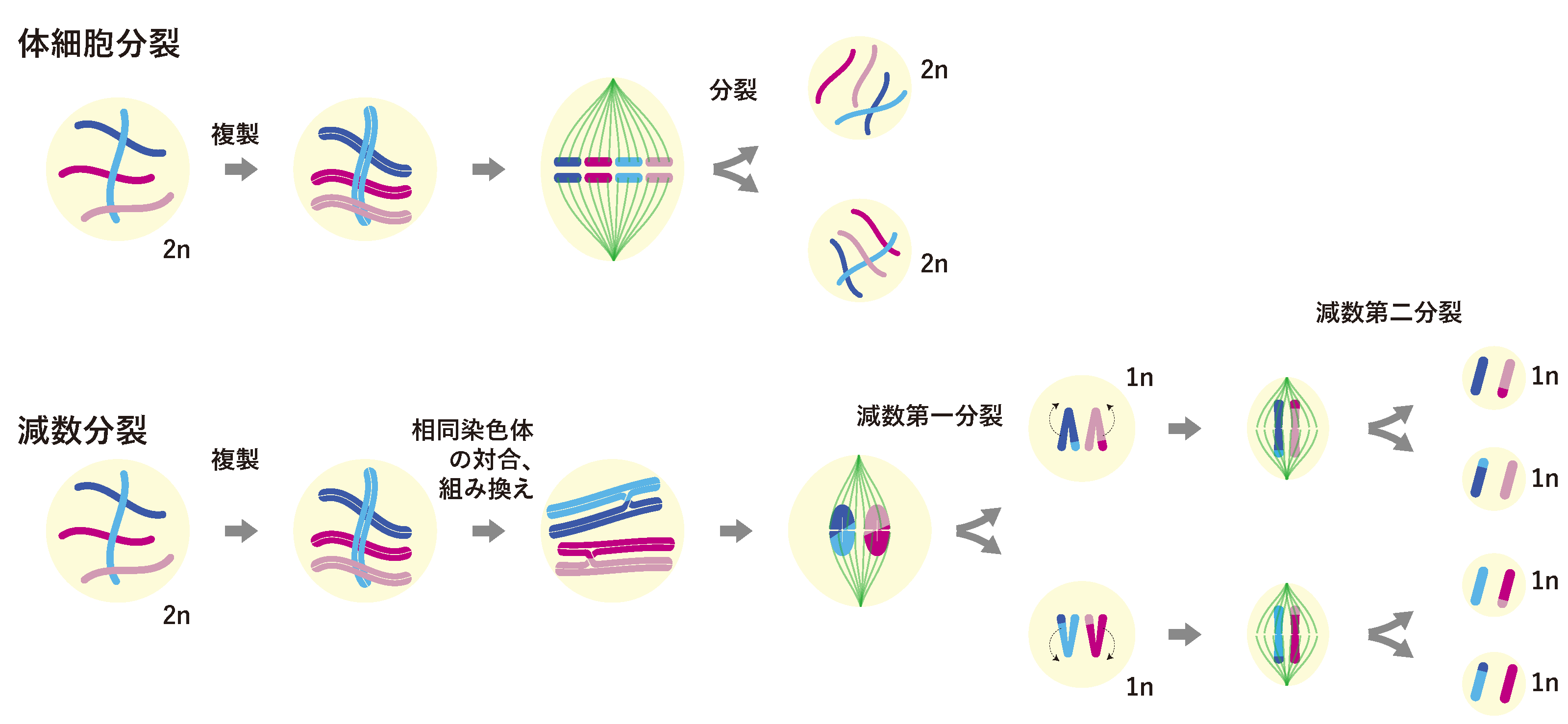
体細胞分裂では、それぞれの染色体が、複製され、姉妹染色体が分離されるだけです。一方、減数分裂では、普段は独立に存在している相同染色体(例えば、母方第一染色体と、父方第一染色体)が、複製されたあとに、2回連続して分離されることによって半数化されます。これは減数第一分裂、第二分裂と呼ばれます(図1)。このため生殖細胞は、第一分裂において分離する染色体の領域、第二分裂において分離する領域を、分裂前にあらかじめ決めておかなければなりません。
私たちヒトでは、染色体上のセントロメアと呼ばれる特定のDNA領域を用いて、減数分裂における染色体分離面を決めており、第一分裂では、セントロメア以外の領域において染色体を分離し、第二分裂では、セントロメア領域において染色体を分離することが知られています。
一方、線虫や一部の植物や昆虫では、このセントロメア様の働きをするDNA領域が染色体全体に広がっており、ヒトと同じように、セントロメアとそれ以外、の2つの領域に染色体を分けることができません。では、線虫では、どのように染色体を2つの領域に分けているのでしょうか?
長さを比べることで染色体を分離する線虫の減数分裂
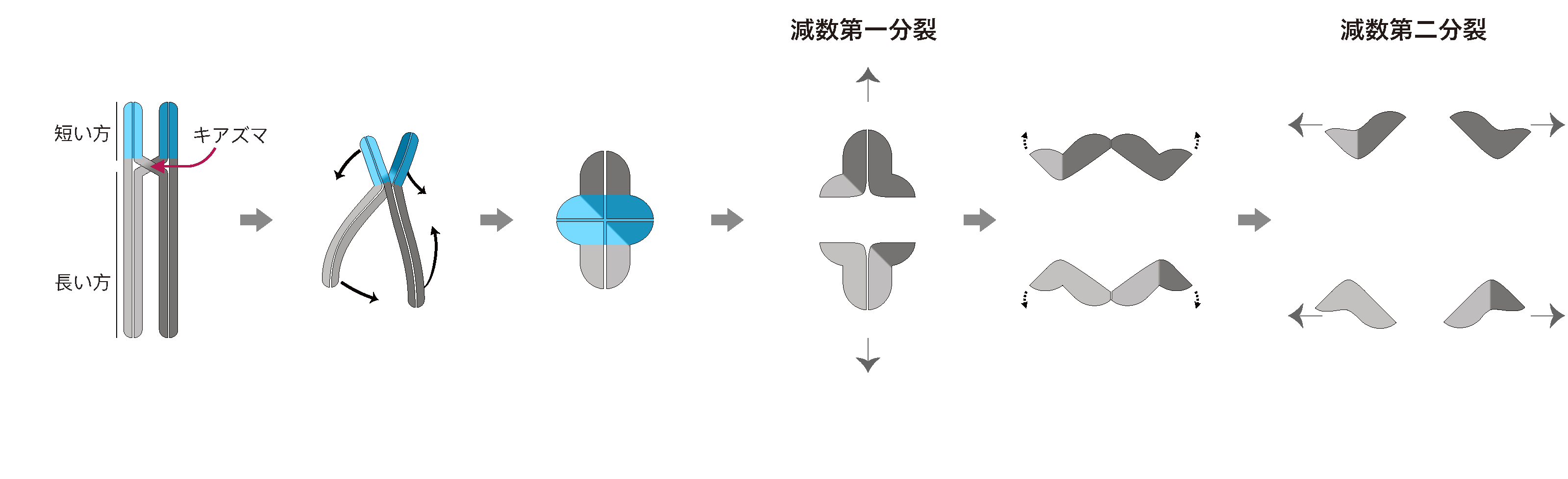
減数分裂では、減数分裂前期と呼ばれる時期に、母方から受け継いだ染色体と、父方から受け継いだ染色体がペアになり、さらに母方と父方の染色体を一部切断して、つなぎかえられることで、母由来と父由来の遺伝子の組み合わせがシャッフルされます。この染色体がつなぎ変えられた部分は“キアズマ”と呼ばれ、線虫では、相同染色体のペアにつき、必ず1つのキアズマが作られます。
実は、線虫の生殖細胞は、キアズマから染色体末端までの距離を比べることで、染色体末端まで短い方と、長い方、という分け方で染色体を2つの領域に分けることが知られています(図2)。
減数第一分裂では、短い方の染色体領域において染色体が分離され、第二分裂では長い方の染色体領域において分離されます。キアズマはいつも決まった場所に作られるのではなく、染色体上の色々な場所に作られるため、線虫の細胞は、キアズマが作られた後に、染色体のどちら側が短くて、どちら側が長いかな?ということを、定規も使わず、目で見ることもなく、検出できているということが、この現象からわかります。
細胞は、生体分子の様々な物理化学的性質を駆使して、細胞内の器官や複合体のサイズを検出したり、調節できることが知られていますが、定規がなくても、2つのものの長さを比べることだってできる!ということを、線虫の細胞は教えてくれます。
参考資料
1) Hillers K.J., Jantsch V., Martinez-Perez E., Yanowitz J.L. Meiosis. (May 4, 2017), WormBook, ed. The C. elegans Research Community, doi/10.1895/wormbook.1.178.1, http://www.wormbook.org.
監修者
カールトン ピーター
アメリカ合衆国、シアトル生まれ。カリフォルニア大学バークレー校博士課程修了。現在、京都大学、大学院生命科学研究科の准教授として減数分裂に関する研究を行なっている。自身が担当する線虫の実習授業では、学生たちと様々な場所から野生の線虫株を採集、同定しており、いつか新種の線虫を発見することを目指している。
佐藤 綾
日本、大阪生まれ。カリフォルニア大学バークレー校博士課程修了。現在、京都大学、大学院生命科学研究科カールトン研究室の博士研究員として減数分裂に関する研究を行なっている。
研究室HP
https://www.carltonlab.org/
©2026 虫の集い(線虫研究者コミュニティ)「1. 減数分裂(生殖腺で精子と卵子ができるまで)」CC BY
2. 受精 ~生命誕生の瞬間~
どうやって線虫の子供ができるのか?
性をもつ生物にとって、受精とは生命誕生の瞬間です。親由来の卵と精子が融合して受精卵となり、このたったひとつの細胞が分裂を繰り返すことで動物のからだが形づくられていきます。大人になれば、また卵や精子をつくり、その中に含まれる遺伝子が次の世代へと受け継がれていきます。このように受精は生命をつなぐとても神秘的な現象です。
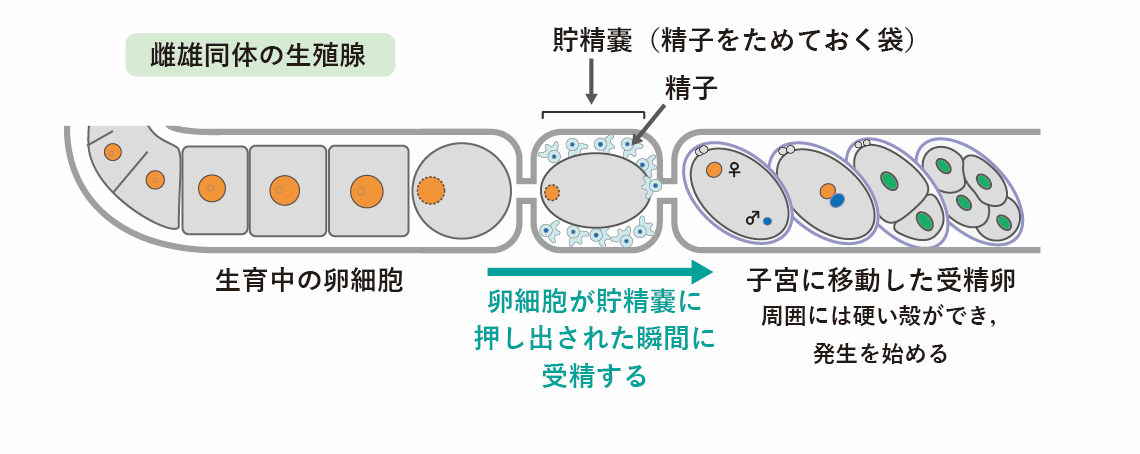
線虫C. elegansには「雌雄同体」と「雄」という2つの性別があります。雌雄同体は自分自身で卵と精子の両方を作り、それらが受精して子供になります。雌雄同体の卵と雄の精子が受精する場合もあります。
卵や精子は、からだの中にある生殖腺とよばれる細長いチューブのような場所で作られます(図1)。雌雄同体の体内では、精子は「貯精嚢」という場所に保管されています。生殖腺で作られた卵は、たくさんの精子が入った貯精嚢に向けて押し出され、それと同時にただひとつの精子と受精します。受精卵はその後、貯精嚢から隣の子宮へ押し出され、しばらくすると自身の周囲に硬い殻(卵殻)を作ります。受精して約1時間後には卵殻の中で最初の細胞分裂が始まり、さらに分裂を繰り返しながらだんだんと線虫のかたちができてきます。この間に受精卵は体外に産卵され、受精してから約14時間後には小さな幼虫が卵の殻から出てきます。
線虫を使って生命誕生の謎に迫る
卵と精子は遺伝子(DNA)を子孫に運ぶための特別な細胞です。それらが受精することによって、あらゆる種類の細胞になることができる能力(全能性)を持った細胞へと変化します。線虫は体が透明なので、卵や精子が作られる時期から受精の瞬間、さらに受精卵の中で何が起こっているのかを、生きたまま顕微鏡で観察することができます。この特徴を活かすことによって、受精卵の中で繰り広げられる生命誕生の秘密が次々と解き明かされています。
たとえば、受精卵の中では、卵や精子に由来する不要となった成分を分解しつつも、その素材をリサイクルすることによって、その後の発生に必要な成分へと置き換える、いわゆるスクラップ・アンド・ビルドが起きていることがわかってきました。このような現象は多くの動物の受精卵に共通することもわかってきています。
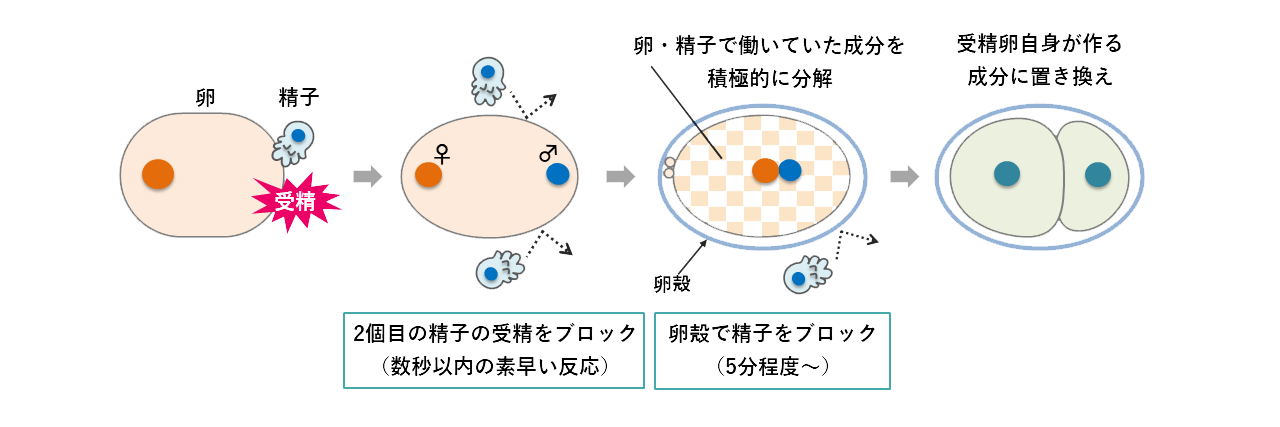
不思議なことに、線虫やヒトの卵はたくさんの精子に囲まれても必ずひとつの精子とのみ受精します。2つ以上の精子と受精(多精)すると雄由来の遺伝子が多くなってしまうので、これを防ぐしくみ(多精拒否)が備わっているのです。線虫では受精後5分ぐらいでできる卵の殻が、2つ目の精子の受精を防ぐことが知られています。また最近の研究から、ひとつ目の精子が受精してから数秒以内に働くすばやい多精拒否のしくみもあることがわかってきました。このように線虫の卵は受精直後と卵殻形成時の2段階の多精拒否のしくみにより、ただひとつの精子とのみ受精するようになっています(図2)。
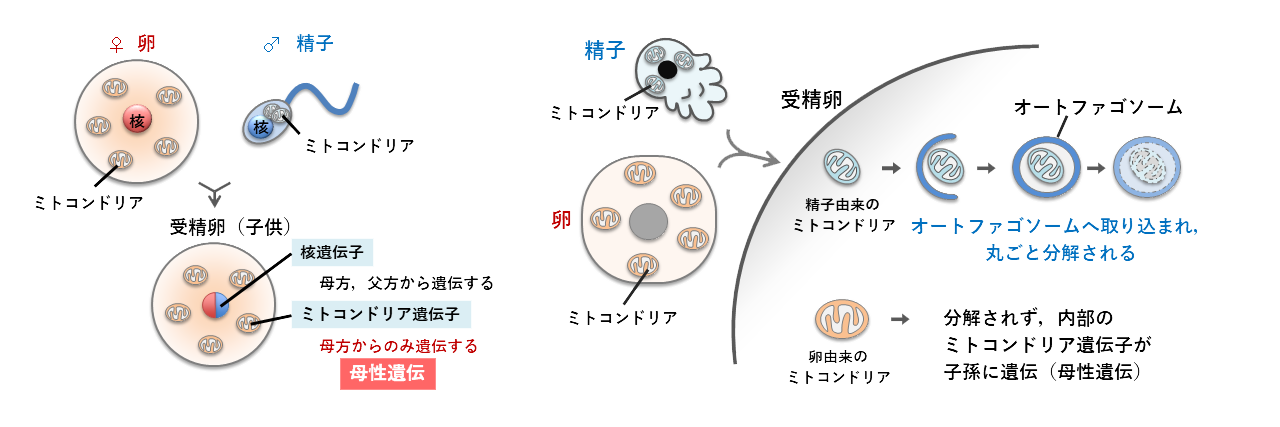
ミトコンドリア遺伝子は母性遺伝する
遺伝子は細胞の中の「核」に保管されていていますが、核以外にも遺伝子が存在する場所があります。それがミトコンドリアの中です。ミトコンドリアは細胞の中にある細胞小器官の1種で、エネルギーを作る発電所のような役割をしています。ミトコンドリアは進化の過程で細菌が細胞の中に住み着いた(共生する)ことで生まれたと考えられており、そのため細菌に由来する独自の遺伝子(DNA)を持つのです。
核の遺伝子は母親(卵)と父親(精子)から半分ずつ子孫へ伝わります。不思議なことにミトコンドリア遺伝子は多くの生物で母親からのみ子孫に伝わるので、このような遺伝のしかたは「母性遺伝」と呼ばれています(図3)。線虫でも雌雄同体の卵と雄の精子が受精した場合はミトコンドリア遺伝子が母性遺伝します。そのしくみや理由は謎につつまれていましたが、線虫の研究から「オートファジー」というしくみが働いていることがわかってきました。
受精すると精子のミトコンドリアは卵の中に入ってきますが、しばらくすると精子のミトコンドリアの周囲にオートファゴソームという袋のような構造ができてきて、中身を丸ごと分解してしまうのです(図3)。では、どうして卵のミトコンドリアは分解されず、精子のミトコンドリアだけ分解されるのでしょうか?それについては現在も引き続き研究が進められています。また、面白いことにヒトの細胞に感染した細菌も精子のミトコンドリアと同じようにオートファジーで分解されることもわかってきています。
参考資料
1) Hall, D.H., Herndon, L.A. and Altun, Z.著 「Introduction to C. elegans Embryo Anatomy」2017年 In WormAtlas. doi:10.3908/wormatlas.4.1
2) 大島靖美 著.「線虫の研究とノーベル賞への道-1ミリの虫の研究がなぜ3度ノーベル賞を受賞したか-」2015年 裳華房 ISBN 978-4-7853-5863-1.
3) 「多精子受精拒否の仕組みの一端を解明 ~なぜ卵母細胞(卵子)はただ1つの精子とのみ受精するのか~」2024年 群馬大学生体調節研究所HP プレスリリース記事 https://www.imcr.gunma-u.ac.jp/cms_imcr/wp-content/uploads/2024/01/dcb83577e9795fdcc9bc08cd5c98b4f1.pdf
4) 佐藤美由紀,佐藤裕公,佐藤健 著.「初期発生におけるリソソーム分解の生理機能と分子メカニズム」2019年 生化学 91(5):643-651. https://seikagaku.jbsoc.or.jp/10.14952/SEIKAGAKU.2019.910643/data/index.html
5) 佐藤美由紀 著 「父由来のミトコンドリアゲノムが消されるしくみ」2015年 JT生命誌研究館 季刊「生命誌」85号. https://www.brh.co.jp/publication/journal/085/research/2
監修者
佐藤美由紀
岐阜県生まれ。東京大学大学院博士課程修了。現在は群馬大学・生体調節研究所教授として、ミトコンドリア遺伝子の母性遺伝や生殖細胞でのオートファジーの役割について研究。
佐藤健
大分県生まれ。東京大学大学院博士課程修了。現在、群馬大学・生体調節研究所・細胞構造分野教授として、線虫やマウスを用いて動物が受精して発生する際に繰り広げられる細胞内リモデリングの現象について研究している。日本生化学会理事なども務める。
研究室HP
http://makukinou.showa.gunma-u.ac.jp/index.html
http://traffic.dept.med.gunma-u.ac.jp/
©2026 佐藤美由紀・佐藤健「2. 受精 ~生命誕生の瞬間~」CC BY SA
3. 線虫が卵から体の形を作る初期の過程―流れ、回転、めり込みが鍵となるメカニズム
初期発生とは?線虫を使った初期発生研究がなぜ重要?
個体発生とは、生物の個体が受精卵のような未熟な状態から成体になるまでの過程のことです。その中でも初期に起こる一連のイベントが初期発生です。この過程では1)体の前後、背腹、左右の方向を決定したり、2)様々な種類の細胞タイプを作ったり、3)その細胞たちを正しく配置したり、する必要があります。
線虫の初期胚は観察や操作がしやすく、また、初期発生のイベントに異常がある変異体を得やすいという重要な利点があります。この変異体のDNAの中でどの遺伝子が変異しているかを調べることによって、初期発生を支配する遺伝子群(後述するa-PAR, p-PAR)などが初めて同定されました。このような遺伝子はヒトでも保存されています。
最近でも、後述する運命決定因子の一つである生殖顆粒が、水の中にある油滴のように“分離”することで形成されることが、初めて線虫で発見されました。この相分離現象はのちにさまざまな生命現象で確認され、近年の細胞生物学の中で一大潮流(いわゆるバズりです)を起こしています。
このように、線虫は一見単純な構造をしていますが、研究者たちが新しい因子を“見つける”ことができ、 “深く”仕組みを理解し、革新的なアイデアを“試す”ことがしやすいので、研究界においてしばしばトレンドリーダーになることがあるのです。たまに間違った仮説が横行することもありますが、巨視的に見ると科学界は建設的な批判の応酬による自己修復能力を持っているので、間違った仮説は忘れられていきます。
体の前後、背腹、左右軸ができるきっかけ
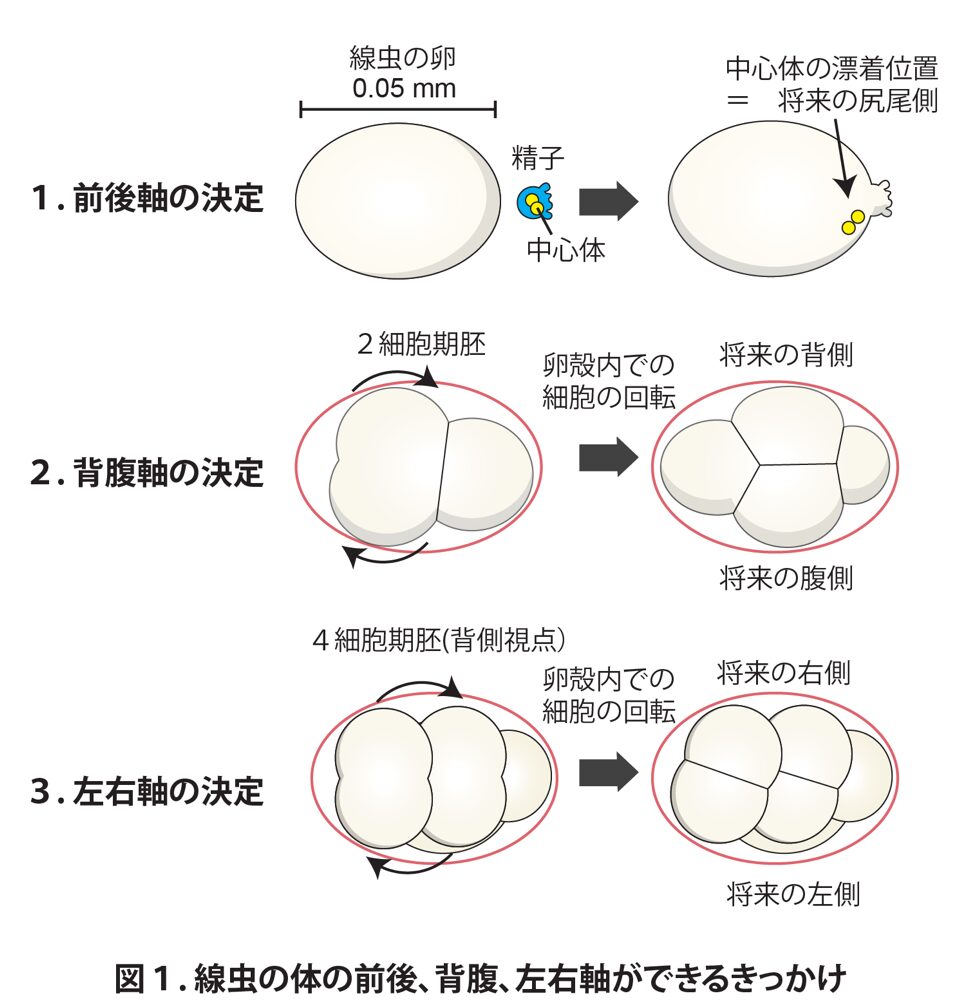
ほぼ全ての動物の初期発生では、まずからだの前後を決める必要があります。前後軸は進化的に最も古くからある体軸であり、動物がえさを探すときに動きまわるためにも、将来からだのどちら側が口になるか肛門になるかを決めるためにも、クラゲのような単純な構造の動物からヒトに至るまで非常に重要なものです。線虫では、精子が卵子に入り、精子に含まれる中心体というタンパク質でできた物体が流れて漂着する場所が体の後ろ側、つまり将来尻尾になるように決まります(図1上段)。
クラゲには背腹の違いはありませんが、線虫を含む、より複雑な動物では、背側と腹側とでからだの機能が違っています。線虫では、2細胞期胚の細胞が分裂するときに卵殻内で回転する方向によって背側と腹側が決まります(図1中段)。前後と背腹が決まると、幾何的に自ずと左右軸も決まります。
外側から見ると、ヒトを含む多くの動物の左右は対称な構造ですが、からだの中の臓器の配置や形は非対称になっています(左右差と呼びます)。線虫の左右差は、4細胞期胚の細胞が分裂するときに回転することで生まれます(図1下段)。これらの回転が重要であることは、その向きを実験者が逆にすることで背腹や左右の臓器の配置が逆転した個体が生まれることにより示されました。
色々な細胞ができる仕組み
ではなぜこのような比較的単純なしくみ(流れ、回転)が前後、背腹、左右の差を生み出すかというと、体軸に沿って細胞の種類に違いが生まれるからです。
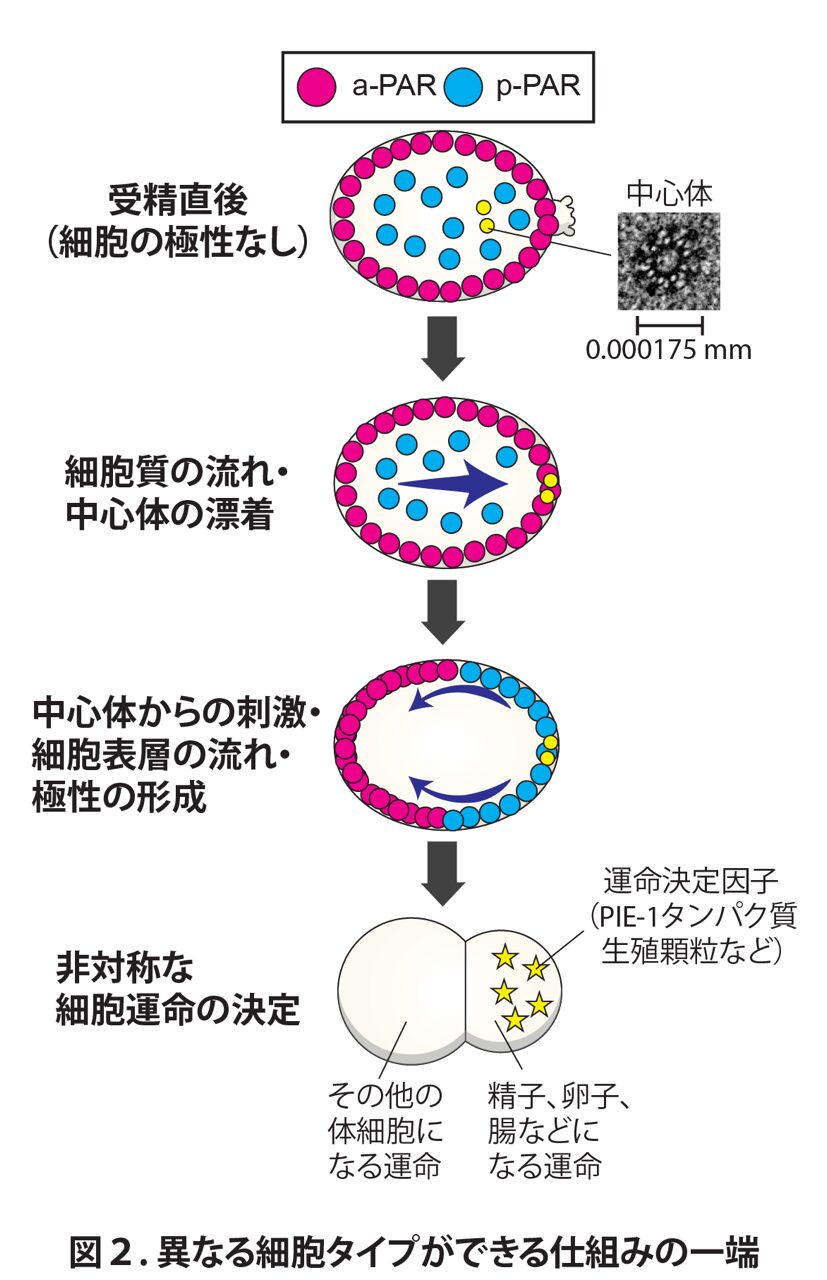
前後軸形成時の例を用いて説明します。上で述べたように、体の前後軸は精子に含まれる中心体が流れて漂着する場所で決まります(図1上段)。これは中心体という細胞小器官に含まれるタンパク質が受精卵の中にあるタンパク質に影響をもたらすからです(図2)。
具体的にいうと、a-PARというタンパク質複合体とp-PARというタンパク質複合体があり、受精前はそれらが前後の差なく一様に分布していますが(図2上2段)、中心体由来の刺激により細胞の表層の流れが発生してa-PARが前側に運ばれ、さらに別のしくみの助けもありp-PARが後ろ側の表層に局在するようになります。この非対称な局在が、さらに下流の運命決定因子と言われるタンパク質の局在を非対称に制御します。これにより、受精卵の最初の卵割後に前側の細胞は体細胞に、後ろ側の細胞は生殖細胞を含む細胞になる“運命”が決定されます。
体の形づくりの仕組み
いろいろな細胞ができただけではからだを形作るには不十分です。それらが形状や配置を変えていく必要があります。その代表的な例が原腸陥入です。20世紀を代表する著名な発生生物学者であるウォルパートは、「人生の中で最も重要な瞬間は冠婚葬祭ではなく原腸陥入である」とかつて言いました。
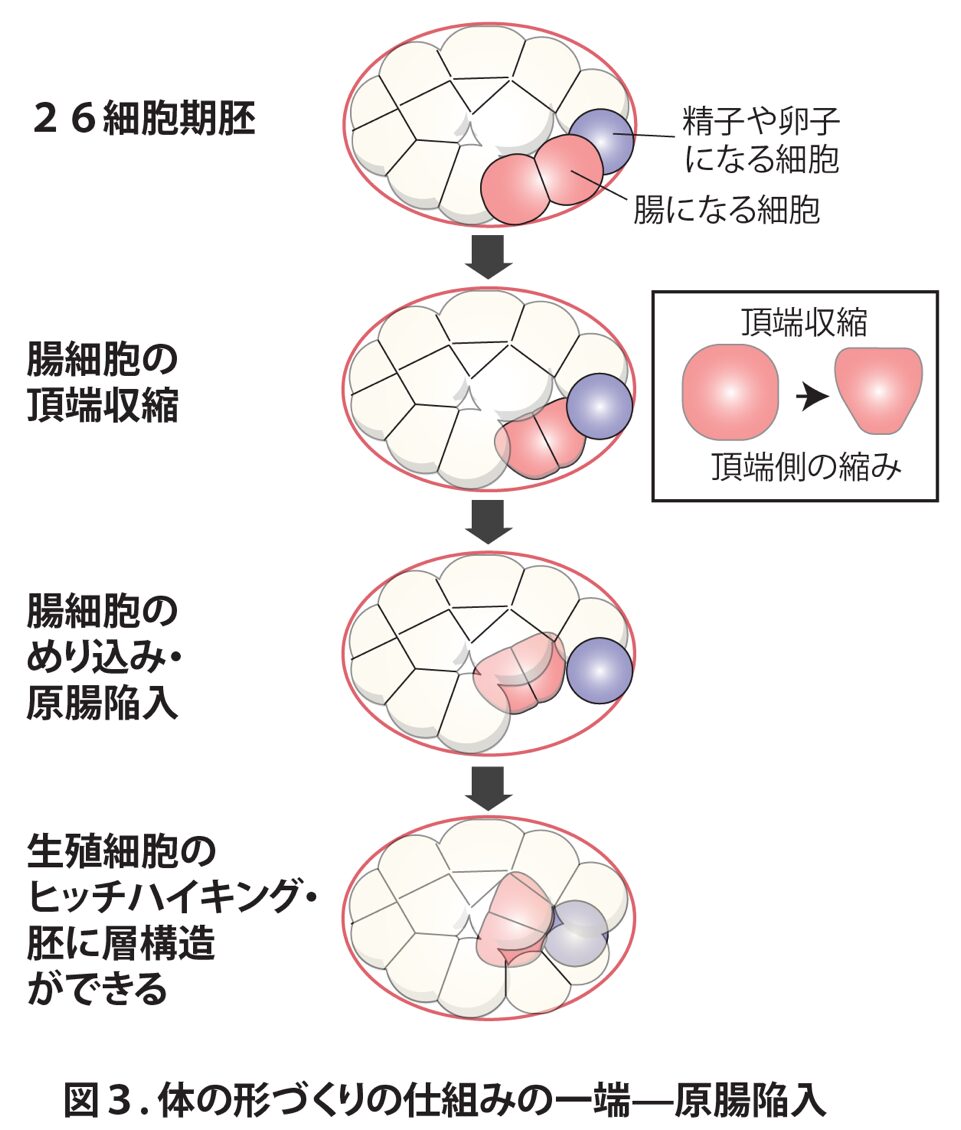
線虫の原腸陥入は比較的単純ですが、より複雑な生き物同様、頂端収縮という細胞の形の変化を伴います。この頂端収縮は26細胞期胚で起こります。この時点では胚は細胞群のただの集まりであり、そこに形態的な複雑さはほぼありません。頂端収縮が起こると、腸細胞の外側の表面が縮まることで、腸細胞が胚の内側にめり込んでいきます(図3)。これに伴って将来卵子や精子になる生殖細胞も折り紙のように胚の内側にめり込んでいく一方、表皮になるような細胞は胚の外側にとどまります。
このように、原腸陥入ではからだの中に機能の異なる細胞による層構造ができるのです。原腸陥入ののちにさらに複雑な形づくりも起こりますが、これらも細胞の物理学的な性質(形、動き、硬さなど)を体軸に沿って変えることでコントロールされています。
参考資料
1)Jonathan Slack著 大隈典子 訳 「エッセンシャル発生生物学」, 2002年 羊土社
2)Gan WJ and Motegi F (2021) Mechanochemical Control of Symmetry Breaking in the Caenorhabditis elegans Zygote. Front. Cell Dev. Biol. 8:619869. doi: 10.3389/fcell.2020.619869
3) Seirin-Lee S., Yamamoto, K., and Kumura A. (2022) The extra-embryonic space and the local contour are crucial geometric constraints regulating cell arrangement Development. 149(9):dev200401.
監修者
杉岡 賢史
ブリティシュコロンビア大学で研究室を主宰。線虫を用いて初期胚発生の研究を行っている。
研究室HP
https://www.zoology.ubc.ca/~sugioka/
©2026 杉岡賢史「3. 線虫が卵から体の形を作る初期の過程―流れ、回転、めり込みが鍵となるメカニズム」CC BY
4. 細胞系譜(体細胞の運命決定)
はじめに
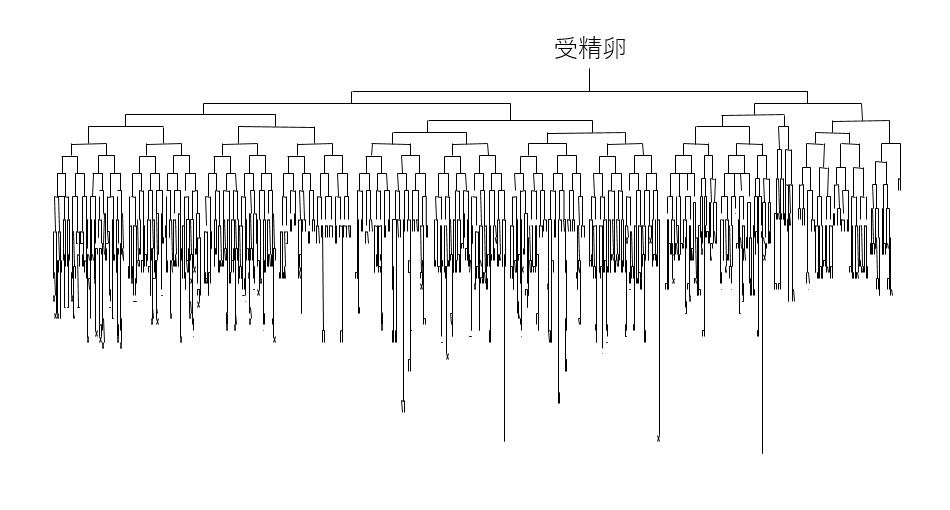
線虫の受精卵から成虫までの細胞分裂そして分化のパターン(細胞系譜)は、John SulstonとRobert Horvitzらによって、直接観察により決定されました (図1は胚発生時の細胞系譜)。驚くことに細胞系譜には個体差がほとんどありません。細胞が死ぬという現象でさえも、特定の細胞が生まれた直後に死ぬことが明らかにされ、プログラム細胞死と名づけられました。細胞系譜と細胞死のメカニズムの発見に対し、2003年にノーベル生理学・医学賞が授与されています(受賞者はSulston、Horvitzおよび研究を主導したSydney Brenner)。細胞系譜に個体差がないのは、分裂や分化が非常に厳密に制御されているからです。細胞系譜がどのように制御されているのか、紹介します。
動物の体には様々な種類の細胞が存在します。発生の際に、受精卵というひとつの細胞が分裂することで、多種多様な細胞が作られますが、分裂により生じた細胞(娘細胞)が全く同じであれば、細胞の数は増えても種類は増えません。細胞の種類を増やすメカニズムは大きく分けて二種類あります。一つは、細胞が外部の環境(隣接細胞との接触や、シグナル分子の受容など)が娘細胞間で異なるため、娘細胞が異なる性質(細胞運命と呼ぶ)を獲得します(機構①)。もう一つは、娘細胞が生まれるときに、異なる物質を母細胞から受け継ぐために、異なる細胞運命を獲得する場合で、非対称分裂と呼ばれます(機構②)。線虫の発生ではこれを組み合わせて多様な細胞を作っていきます。
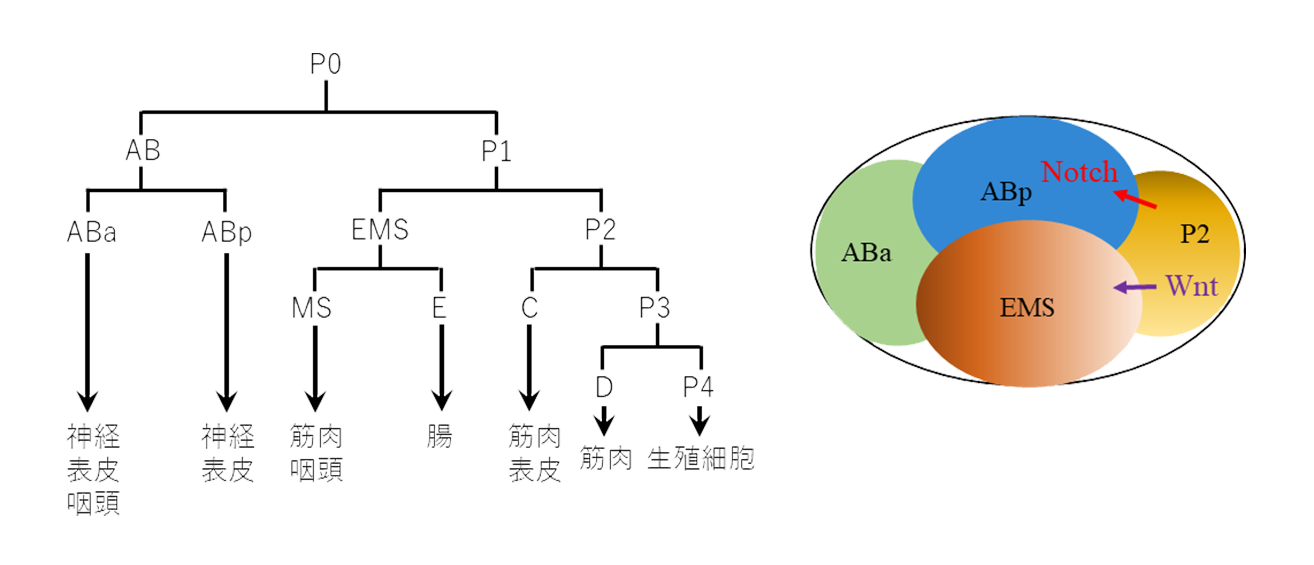
左:発生初期の細胞系譜(細胞分裂パターンと細胞運命)。分裂ごとに娘細胞は異なる運命を獲得する。それぞれの細胞がその後作り出す主な細胞種を示します。
右:4細胞期の細胞間シグナル。P2からABpへのシグナルはNotch受容体を介して(赤)、ABpの運命の違いを作り出します(機構①)。P2からEMSへのWntシグナル(紫)は、EMS細胞内に極性を作り出し、その娘細胞が異なる運命になります(機構②)。
胚発生の細胞系譜制御
項目「初期発生」で触れたように、受精卵(P0細胞)の初めの分裂は非対称で(機構②)、体細胞を作るABと生殖細胞などを作るP1細胞ができます(図2左)。P1およびその子孫の生殖細胞を作る細胞(P系列細胞)は、同じ機構により、非対称分裂を繰り返し、体細胞(EMS細胞など)と生殖細胞を作り出します。AB細胞の娘細胞は、生まれたときには同じ性質ですが、ABp細胞だけが、隣接細胞P2からのシグナル(Notch受容体のリガンド)を受容するため(機構①)、ABa細胞と異なる運命を獲得します(図2右)。次にEMS細胞が分裂し、筋肉などを作るMS細胞と、腸を作るE細胞が作られます。MSとE細胞は生まれたときから異なっており(機構②)、ばらばらにしても、それぞれ筋肉と腸を作ります。この非対称分裂は、Wntシグナル伝達経路の働きにより、分裂後、シグナル分子βカテニンや転写因子TCFの核内濃度が娘細胞間で異なっており、これにより娘細胞は異なる細胞になります。この非対称性は細胞外のシグナル分子Wntによって制御されていますが、このシグナルは機構①のように娘細胞に働いて、違いを誘導するのではなく、母細胞に働いて、その内部にシグナル分子の偏り(極性)を作ることで、娘細胞の違いを作ります(図2右)。
その後、ほとんどの体細胞の分裂でTCF転写因子の非対称核局在が観察され、非対称分裂(機構②)を繰り返すことで、異なる娘細胞が作られていきます。加えて、いくつかの娘細胞に、ABp細胞運命を制御したのと同様、隣接細胞との接触を介したNotchシグナルが働き、特異的な細胞運命を誘導します(機構①)。このように、体細胞の多様な運命は機構①と②の組み合わせで決定されていきます。
幼虫の細胞系譜制御
卵から孵化した幼虫は生存に必要な細胞はすでに持っています。幼虫期では、体の成長に伴う細胞の補充と生殖器官の形成に必要な細胞分裂が起こります。これらの細胞分裂もほとんどがWntシグナルに制御された非対称分裂(機構②)で、娘細胞は異なる運命を獲得します。一方、細胞外シグナルによる細胞運命制御(機構①)も働いており、その代表的な陰門形成について紹介します(図3)。陰門(産卵口)は、L3幼虫期に6つの前駆細胞のうち3つ(P5.p、P6.p、P7.p)から作られます。P6.pに近接するアンカー細胞から分泌されるシグナル分子によって、細胞内でRas-Mapキナーゼ経路が活性化され、アンカー細胞に最も近接するP6.p細胞は1゜、少し離れたP5.pとP7.p細胞は2゜と呼ばれる細胞運命を獲得します。加えて。1゜と2゜の違いには、近接細胞間のNotch受容体を介したシグナルも関与しており、この二種のシグナル機構の組み合わせで、2゜1゜2゜という鏡像対称な細胞系譜、そして鏡像対称な陰門の形態が作られます。
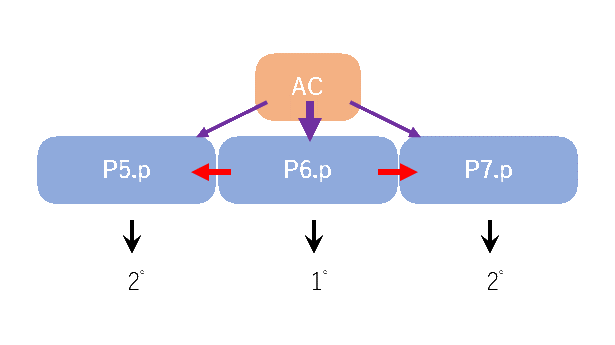
陰門発生の際の細胞間シグナル。アンカー細胞(AC)からのシグナル(紫)を強く受けるP6.p細胞は1゜、弱く受けるP5.p、P7.p細胞が2゜の運命を獲得します。隣接細胞間のNotch受容体を介したシグナル(赤)も2゜運命を誘導します。
終わりに
このように線虫の発生では、機構①②の組み合わせにより、厳密に細胞運命が制御されています。このため、発生の途中の個々の細胞が、どのように生じたのか、これからどのような細胞を作るのかがわかります。様々な変異体や遺伝子操作で、運命に変化が起これば、その原因を遡って調べることができます。そして、異常が起こる際の、遺伝子発現、分子の機能・局在の変化を調べることで、どのように細胞運命・細胞系譜が決められるのかを知ることができます。
監修者
澤 斉
大阪生まれ。京都大学大学院博士課程修了。現在、遺伝学研究所教授として「発生の仕組み」の研究・講義を担当している。日本発生生物学会理事なども務めた。
研究室HP
http://square.umin.ac.jp/Nemalab/Home.html
©2026 虫の集い(線虫研究者コミュニティ)「4. 細胞系譜(体細胞の運命決定)」CC BY
5. 形態形成・器官形成
C. elegansのからだの一番表面は外骨格であるキューティクル(細胞外マトリックス)があり、その内側に表皮と体壁筋の層があります。ほとんどの神経細胞は表皮の内面に付着して存在しています。さらにその内側は偽体腔と呼ばれており、ここには咽頭、消化管、生殖巣の3種類の器官が存在しています。咽頭はヒトの場合の口と喉、消化管は胃や腸、生殖巣は卵巣や精巣にあたります。動物の多くの器官は上皮と呼ばれる一層の細胞層に覆われており、さらに上皮の表面は基底膜と呼ばれる細胞外マトリックスに覆われています。動物の発生過程では様々な組織・器官の形態形成が進行します。形態形成の主な要因となるのは細胞の分裂、分化、移動、形態変化、基底膜の形成、細胞間接着の変化、細胞基底膜間接着の変化などがあげられます。ここでは生殖巣の形成を中心として、器官形成のしくみを解説します。
生殖巣の形成
C. elegansはからだの中心から前後にU字型に伸びたチューブ状の生殖巣を持っています。孵化直後の1齢幼虫の腹部中央に、前後方向に4個の細胞が並び基底膜で覆われた生殖巣原基が存在します。このうち、中央の2個の細胞Z3とZ3は始原生殖細胞で、分裂を繰り返し、成虫期には精子や卵に分化します。両端の2個の細胞Z1とZ4は同じ決まったパターンで分裂を行い、それぞれが前側と後ろ側の生殖巣の鞘をつくる10個の細胞と1個の先端細胞DTCに分化します。DTCは1齢幼虫期にZ1とZ4の2回目の分裂で生じ、4齢幼虫期までの間にU字型の移動を行うことにより、U字型の生殖巣の形成の指揮をとる働きをします(図1)。このようなチューブ状の器官は線虫独特のものではなく、哺乳類で言えば、肺や腎臓、肝臓、乳腺、唾液腺など多くの器官は複雑に枝分かれした上皮チューブでできています。これらの器官は哺乳類の発生の過程で、単純なチューブが伸長と枝分かれを繰り返すことにより形成されます。
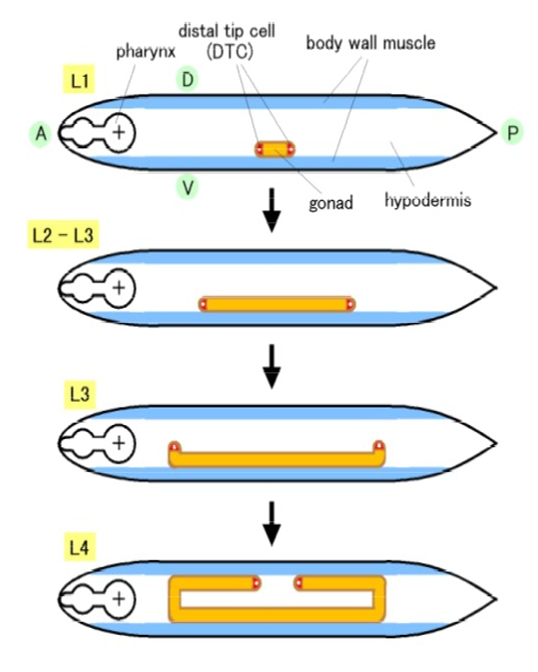
先端細胞の移動のしくみ
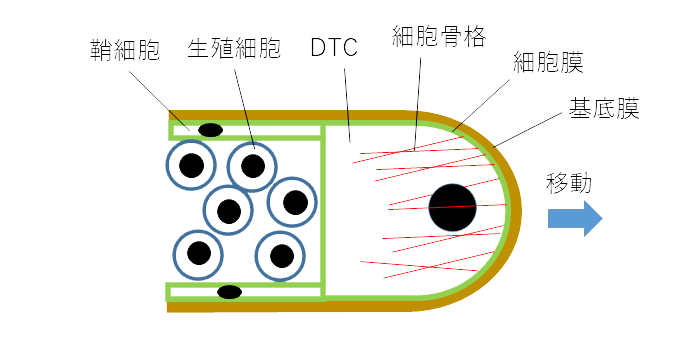
ではDTCはどのようにして移動を行うのでしょうか。上に述べたように生殖巣の表面は基底膜で覆われています。DTCが移動を行う時には、DTCは基底膜に接着しており、これをアクチンフィラメントなどの細胞骨格の力で押し広げるようにして移動していきます。この時、生殖巣の先端部分には新しい基底膜がどんどん付加されていきます。基底膜の主成分はコラーゲンと呼ばれるタンパク質です(図2)。
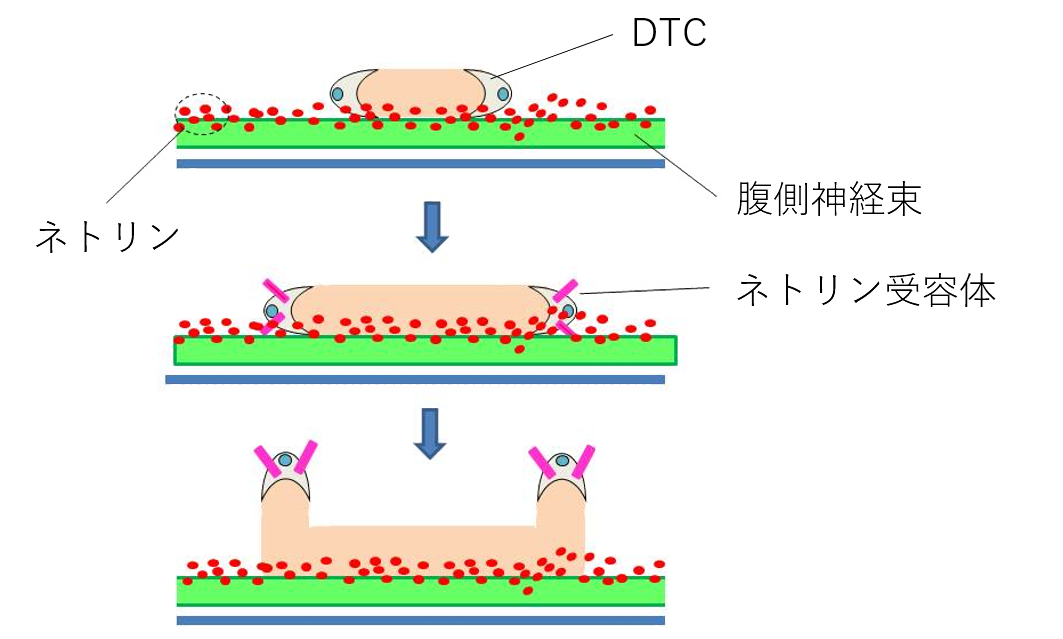
DTCの移動により、生殖巣は初め腹側の体壁筋に沿って前後方向に伸長します。DTCは2回の方向転換をして、U字型の生殖巣の形成を指揮しますが、この方向転換には細胞移動のガイドと呼ばれるしくみが働きます。線虫の腹側神経束からはネトリンと呼ばれるガイドタンパク質が分泌され、腹側で濃度が高く、背側にいくほど低くなる濃度勾配を形成しています(図3)。3齢幼虫期に達した段階で、DTCは1回目の背側への方向転換をしますが、DTCは方向転換の直前に細胞膜上にネトリンの濃度を感じるアンテナである受容体を発現します。DTCはネトリンの濃度を感じて、より濃度の低い背側に方向転換します。2回目の方向転換にはウイントと呼ばれるガイドタンパク質が働きます。ネトリンやウイントなどのガイドタンパク質は哺乳類の器官形成でも働くことが知られています。
参考資料
E. Jane Albert Hubbard and David Greenstein (2000). The Caenorhabditis elegans Gonad: A Test Tube for Cell and Developmental Biology. Developmental Dynamics 218:2-22.
監修者
西脇 清二
1956年大阪生まれ。線虫の胚発生研究により大阪市立大学より博士学位を取得。NEC基礎研究所、理化学研究所を経て、現在、関西学院大学教授として「線虫の器官形成機構」を研究している。
研究室HP
https://sci-tech.ksc.kwansei.ac.jp/~nishiwaki/
©2026 虫の集い(線虫研究者コミュニティ)「5. 形態形成・器官形成」CC BY
6. プログラム細胞死(アポトーシス)
プログラム細胞死(アポトーシス)は、あらかじめ決められたプログラムにしたがって細胞が死ぬ現象のことです。これは生物が正常に発生し、からだの機能を維持するうえで重要なしくみの一つです。線虫C. elegansの細胞系譜には個体差がなく、雌雄同体の発生過程では1090個の細胞が作られ、そのうち131個の細胞がプログラム細胞死を遂げます。
細胞死が起こらなくなったり、逆に細胞死が過剰に起きるようになった変異体の遺伝学的解析によって、プログラム細胞死に関与する遺伝子群が特定され、その遺伝子経路が明らかになりました。さらに、この遺伝子経路は、線虫からヒトに至るまで多様な生物において保存されていることがわかっています。
細胞死は、死ぬ細胞が決められる「指定段階」、細胞死のプログラムが作動する「殺害段階」、細胞が分解され、近隣の細胞に取り込まれる「実行段階」、の3つのステップで行われます。
「殺害段階」は、EGL-1, CED-9, CED-4が関与する一連のタンパク相互作用によって細胞死プロテアーゼCED-3が活性化されると始まります。
C. elegansの細胞系譜の決定とプログラム細胞死の遺伝的経路の解明で主導的な役割を果たしたシドニー・ブレナー、ジョン・サルストン、H.ロバート・ホロヴィッツには2002年のノーベル生理学・医学賞が授与されました。
参考資料
1) Conradt B, Xue D. Programmed cell death, WormBook. 2005 Oct 6:1-13. doi: 10.1895/wormbook.1.32.1.
2) Metzstein M, Stanfield GM, Horvitz HR. Genetics of programmed cell death in C elegans: past, present and future
監修者
杉本 亜砂子
東京都生まれ。東京大学大学院理学系研究科博士課程修了。現在、東北大学 理事・副学長(研究担当)、大学院生命科学研究科・教授。日本分子生物学会理事長、WormBoard(国際線虫会議)初代アジア代表も務めた。
研究室HP
https://devdyn.wixsite.com/sugimoto-lab
©2026 虫の集い(線虫研究者コミュニティ)「6. プログラム細胞死(アポトーシス)」CC BY